
| - Características generales |
Clamidia psittaci
es una de las cuatro especies de Clamidia caracterizadas hasta hoy. Infecta
preferentemente ganado, aves, animales domésticos en general y
al hombre, pero también peces, tortugas y cocodrilos. Una de las
vías de contagio más importantes es el contacto con los aerosoles
resultantes de deposiciones secas de aves o animales de granja, ya que
naturalmente esta especie de Clamidia desarrolla una forma de infección
persistente intestinal en los animales.
A diferencia
de Clamidia trachomatis (cuyo huésped principal es el hombre), la
especie que describiremos posee un tropismo celular muy amplio, pudiendo
desarrollarse en varios tejidos, si bien esta característica no
se extiende a todas sus cepas. Algunos ejemplos incluyen células
epiteliales en mucosas, cloaca (en aves), pulmón, bazo.
Las enfermedades
que es capaz de producir van desde la psittacosis en el hombre a placentopatías
abortivas en ovejas, vesiculitis, encefalomielitis bovina, conjuntivitis,
hepatitis, poliartritis, y otras.
Se describieron
para esta especie, las proteínas de 22 kDa, 34 kDa, y 52 kDa, que
se observan expresadas durante el desarrollo en la célula huésped,
pero no en el cuerpo elemental (CE) y cuya función sería
la de facilitar el ingreso de nutrientes.
La respuesta
del huésped está mediada principalmente por linfocitos T,
tal como ha podido comprobarse en ratones. De hecho en ratones sin timo
que fueron previamente infectados con C. psittaci y curados de su infección,
se observó susceptibilidad ante reinfección con la misma
cepa, (dando infección persistente), no así con ratones LB
deficientes, que fueron, bajo las mismas condiciones, refractarios a la
reinfección. Más aún, se ha logrado brindar inmunidad
pasiva con una infusión en ratones nunca infectados, de linfocitos
T provenientes de ratones que resolvieron su infección.
El mecanismo
queda a cargo del IFN-g
el TNF-a, que
inducen una infección persistente en macrofagos y en LT impiden
la evasión del lisosoma, dando inclusiones que nunca desarrollan
y cuyo contenido es acídico.
La lisis de
las células infectadas es preferentemente por LT CD8 +, y son más
fácilmente lisables que las infectadas con C. Trachomatis, puesto
que C. psittaci es susceptible de sufrir lisis Ag-independiente.
La presentación
antigénica se realiza predominantemente por MHC clase I, pero cuando
se forma el fagolisosoma en LT, en la membrana de la vacuola se detectan
MHC clase II, lo que lleva a pensar que también ellas podrían
actuar como presentadoras de Ag en ese contexto.
-Ciclo de crecimiento -(comparación con C. trachomatis)-:
Infecta
la célula bajo la forma de CE entrando en contacto con ella preferentemente
en zonas de la membrana cubiertas de clatrina, o en la base de las microvellosidades.
En caso de haber ingresado más de un CE sus inclusiones se fusionan
para formar una sola, y esta fusión no se ve impedida frente a inhibidores
de la polimerización de los microtúbulos, cosa que sí
ocurre en el caso de C. Trachomatis. Se ubica inmediatamente en la zona
perinuclear, en contacto con el aparato de Golgi, de donde tomará
esfingosina para sus esfingolípidos durante su desarrollo y multiplicación
por fisión binaria, una vez que se halle como CR. Esta característica
es compartida con C. trachomatis, no así la morfología de
su inclusión. De hecho, pudo observarse que varias de las cepas
más utilizadas para estudio, como la GPIC (guinea pig inclusion
conjunctivitis) desarrollan inclusiones lobuladas, que previo a
la liberación del CE hacia el final del ciclo, evolucionan a distintas
inclusiones (cada lóbulo origina una), e incluso se disgregan las
membranas, liberando CE al citoplasma, antes de la lisis celular. Esta
liberación del CE al citoplasma no siempre es masiva, y no siempre
es seguida por lisis de la célula huésped, lo cual abre las
puertas hacia una nueva concepción de la persistencia espontánea.
Este último
mecanismo fue descrito para las cepas A y D 92/1293, y resulta coincidente
con una mayor virulencia, más rápido desarrollo, mayor infectividad
y la capacidad de producir mayor número de inclusiones por célula
y más alto porcentaje de CE en cada inclusión. La comparación
es con la cepa B, cuyo ciclo de vida es más lento, tiene tropismo
tisular menos amplio, y produce infecciones que nunca son mortales.
Precisamente
debido a estas diferencias entre cepas, que no permiten generalizar respecto
de la especie, es que se dispone de mucho menos información acerca
de C. psittaci.
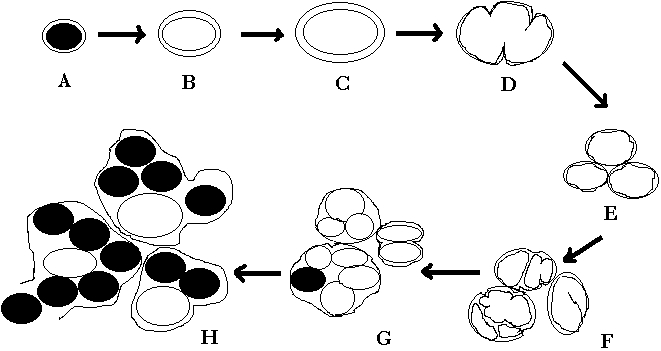 -
Modelo del desarrollo de una inclusión clamidial lobulada
-
Modelo del desarrollo de una inclusión clamidial lobulada
- Por deficiencia
de nutrientes: Si se incuban células no infectadas durante 24
hs., se recoge el sobrenadante de cultivo y se lo utiliza como medio para
el crecimiento de células recién infectadas, se observan
formas aberrantes de desarrollo , que evolucionan rápidamente al
ciclo de vida normal en cuanto se les cambia el medio de cultivo por uno
fresco.
- Inducida por drogas:
* PENICILINA: Previene la fisión binaria, no la diferenciación de CE a CR. Los CR crecen, dando formas aberrantes en las que al microscopio electrónico pueden verse diversos sitios de condensación de ADN, que corresponden a las Clamidias individuales producto de la división del CR que tendrá lugar al quitarle la penicilina al cultivo. En ese caso, la infección sigue su curso normal. En caso de no quitar la penicilina, los CR siguen creciendo hasta provocar la lisis celular, y este es el mecanismo por el cual se puede curar una infección con penicilina en altas dosis y con tratamiento prolongado, ya que no existe reinfección de otras células posterior a esa lisis, porque no hubo desarrollo de CE
* D-CICLOSERINA: así como la penicilina, afecta la síntesis de la pared de peptidoglicano en otras bacterias, y su mecanismo de acción sobre Clamidias aún no ha sido elucidado. La persistencia provocada por ésta transcurre de la misma manera que la de la penicilina, y es reversible si al cultivo se le añade su análogo estructural, la D-Ala.
* AMINOPTERINA: Actúa inhibiendo la dihidrofolato reductasa, e impide el desarrollo por falta de tetrahidrofolato, por lo tanto puede revertirse su efecto agregando un exceso de éste al medio. Se ha probado su inducción de persistencia de C. psittaci en Mc Coy.
* CLORANFENICOL: es un inhibidor específico de la síntesis de proteínas, que a altas dosis bloquea la diferenciación de CE a CR. Aparentemente los CE son digeridos por la célula pero luego, si se quita el cloranfenicol, se ve nuevamente desarrollo de Clamidias.
* ACIDO NALIDIXICO: inhibe la síntesis de ARN. No limita el pasaje de CE a CR pero inhibe fuertemente la fisión binaria, y es el único que no limita la síntesis de macromoléculas en C. psittaci.
* RIFAMPICINA: inhibe la síntesis de ADN . si se coloca en el cultivo desde sus inicios, previene la diferenciación de CE a CR si se coloca 12 horas post infección, inhibe la multiplicación de los CR
Con cualquiera
de los antibióticos citados, puede recuperarse la capacidad normal
de infección de la Clamidia en cuestión, con solo liberar
el cultivo de la acción de la droga.
Lo que se ha
observado durante la persistencia es un descenso en la expresión
de MOMP, la cual al ser una porina, disminuye la entrada de moléculas
de antibiótico al ser éstas hidrosolubles, y al ser además
antigénica, disminuiría en el huésped la eficacia
de la respuesta inmune.
* IFN-g:
además de su rol en la respuesta inmune, en caso de cultivos en
células epiteliales se ha visto que afecta la enzima que cataliza
el metabolismo de triptofano. El comportamiento de la inclusión
es el mismo que el que se da con penicilina: la endopoligenia que evoluciona
a CR normal ante la quita del agente promotor de la persistencia.
- Persistencia espontánea:
Esta clase de
persistencia no la dan todas las cepas de C. psittaci. Incluso se ha logrado
in vitro con cepas que luego no la demuestran en infecciones naturales.
La describió
inicialmente Moulder en L cells (fibroblastos de ratón) y usando
GPIC.
Este tipo de
infección da lugar a la postulación de la existencia de
una forma críptica de Clamidia, puesto que se cumple cíclicamente
la siguiente sucesión de hechos:
a) La infección
progresa normalmente resultando en la lisis de casi todas las células
del cultivo.
b) Las células
sobrevivientes no evidencian presencia de Clamidias, y son usadas para
un nuevo cultivo.
c) Este nuevo cultivo
se desarrolla con normalidad salvo por la presencia de unas pocas células
visiblemente infectadas.
d) En sucesivos pasajes
no varía la situación hasta que se observa un repentino aumento
de inclusiones clásicas, produciéndose lo que se llamó
wipe out o proceso de limpieza, puesto que esto acaba con la lisis de la
mayoría de las células cultivadas, y estamos otra vez en
a).
Durante los sucesivos
cultivos sin inclusiones visibles, tampoco se detectó ninguna otra
forma de Clamidia (aunque por lo que es de mi conocimiento aún no
se ha intentado rastrear ADN de Clamidia), ni sus CE ni sus Antígenos.
Por ello, sabiendo por la evolución de las células que sí
están infectadas, se recurrió a la "forma críptica"
de Clamidia.
1) Ultraestructural
changes in avian Chlamydia psittaci serovar A B and D-infected Buffalo
Green Monkey cells.
Inf.
Immun. Apr.1996, Vol.64. n 4 p.1265-71.
2) Chlamidia
psittaci infections:a review with emphasis on avian chlamidiosis
Vet.
Micr. 1995 Rev. 45, 93-119.
3) Temporal
analysis of the developing Chlamydia psittaci inclusion by using fluorescence
and electron microscopy.
Inf.
Immun. Oct. 1996, Vol.61, n 10, p. 4269-78.
4)
Distribution of Endosomal. Lysosomal, and Major Histocompatibility complex
markers in a monocytic Cell Line infected with C. psittaci.
Inf.
Immun. Jun 1997. Vol 65. N 6. P. 2437-42.
5) Cross-reactive T-lymphocyte-mediated lysis of C. trachomatis and C. psittaci infected cells. Inf. Immun. Mar.1997, Vol 65, n 3. P.951-6.
6) Murine Granulated metrial gland cells are susceptible to C. psittaci infection in vivo. Inf. Immun. Sept. 1996, Vol. 64,n 9, p.3897-3900.
7) Persistent Chlamidiae: from cell culture to a paradigm for chlamidial pathogenesis. Micr. Rev. Dec.1994, Vol.58. n 4, p.686-699.
8) Effect of penicilin on te multiplication of meningopnemonitis organisms.J. Bact.Oct. 1968, Vol.96. n 4, p. 875-880.
9) Effect of Chloramphenicol, Rifampicin., and Nalidixic Acid on C.psittaci growing in L cells. J.Inf.Dis. Feb.1973 Vol. 127, n 2, p.155-163.
10) Persistent infection of mouse Fibroblasts with C.psittaci: evidence for a cryptic chlamidial form. Inf.Immun. Dec. 1980.Vol.30, n 3. P. 874-883.
11) Attachement defect in mouse fibroblasts persistently infected with C. psittaci. Inf.Immun. Oct. 1981, Vol.34, n 1, p.285-291.
12) Persistent infection el L cells with an ovine abortion strain of C.psittaci. Inf.Immun. Nov 1985, Vl. 50, n 2, p453-458.
13) Lymphokine mediated inhibition o Chamydia replication in mouse fibroblasts is neutralized by anti-gamma interferon inmunoblobulin. Infect.Immun. Vol 42, n 3, p 1152-1159.
14) In vitro activities of Doxycyckine and enrofloxacin against european C.psittaci strains from turkeys. Antim.Ag. Chem. Vol 41 n 12. P. 2800-2801.
15) Cytotoxic
T Lymphocyte mediated Cytolisi of L cells Persistently Infected With Chlamydia
spp.Infec.Immun. Vol 64. N 6.p 1944-1949.
